Výzkum
Úvod do prostorové paměti
 Prostorová navigace potkanů je dokladem existence prostorové paměti,
a je jedním z nejoblíbenějších modelů chování pro studium neurobiologických mechanismů,
které jsou základem učení a paměti, rozhodování, a dalších vyšších kognitivních procesů (O'Keefe a Nadel 1978).
Výhody prostorových úloh jsou mnohé: navigace je přirozená pro většinu druhů,
což usnadňuje komparativní studie (Nekovářová et al 2013). V experimentálních bludištích lze snadno ovládat
dostupné senzorické vstupy (Blahna et al. 2011) a navigační testy mohou být kombinovány s inaktivacemi mozkových okruhů, elektrofyziologií,
nebo pokročilými molekulárními a genetickými technikami. Kombinace těchto přístupů nám může pomoci poodhalit,
jak mozek zpracovává informace a vede k tvorbě chování a akcí.
Prostorová navigace potkanů je dokladem existence prostorové paměti,
a je jedním z nejoblíbenějších modelů chování pro studium neurobiologických mechanismů,
které jsou základem učení a paměti, rozhodování, a dalších vyšších kognitivních procesů (O'Keefe a Nadel 1978).
Výhody prostorových úloh jsou mnohé: navigace je přirozená pro většinu druhů,
což usnadňuje komparativní studie (Nekovářová et al 2013). V experimentálních bludištích lze snadno ovládat
dostupné senzorické vstupy (Blahna et al. 2011) a navigační testy mohou být kombinovány s inaktivacemi mozkových okruhů, elektrofyziologií,
nebo pokročilými molekulárními a genetickými technikami. Kombinace těchto přístupů nám může pomoci poodhalit,
jak mozek zpracovává informace a vede k tvorbě chování a akcí.
Prostorová navigace představuje zvířecí model lidské deklarativní paměti (Eichenbaum 2001)
nebo odlišnou dílčí složkou deklarativní paměti (Morris 2013).
V dynamických prostředích se zvířata a lidé často dostávají do kontaktu s pohybujícími se objekty nebo dokonce celými pohyblivými
prostředími. Prostorová navigace, a to zejména v takovýchto dynamických prostředích, zahrnuje vnímání času (tzv. intervalové časování,
Buhusi a Meck 2005), což může vést k tvorbě časovacích strategií a jejich kombinací s percepcí prostoru (Klement et al 2010).
Obecně se v navigaci v dynamických prostředích uplatňuje více procesů. Kromě reprezentace skrytých cílů (Morris 1981, 1984),
rozpoznání pozice objektů v nepřístupném prostoru (Klement et al. 2010, Levčík et al. 2013a,b),
průběžné aktualizace měnící informace (Morris a Frey, 1997, Telenský et al. 2009, Telenský et al. 2011, Svoboda et al., 2012)
jsou zásadní i behaviorální flexibilita (Burghardt et al. 2012, Lobellová et al., 2013; Hatalová et al., under review)
a kognitivní koordinace více informačních toků (Wesierska et al. 2005, Kubik a Fenton, 2005, Kubík et al., 2014).
Dohromady tyto procesy tvoří vysoce flexibilní a účelný systém prostorového chování na základě
vnitřní reprezentace prostředí ve formě tzv. "kognitivní mapy" (Tolman, 1948).
nahoru
Hipokampus, přilehlé struktury, a jejich role v prostorové paměti
 Hippokampus je fylogeneticky stará mozková kůra, tzv. archikortex. Je propojen v širší funkční síť
sestávající z hippocampus proper (Cornu Ammonis, CA1-4), gyrus dentatus (angl. dentate gyrus; DG), and subikulárního komplexu (Amaral a Witter 1989).
Spolu s okolními oblastmi kůry, především entorhinální, perirhinální a postrhinální kůry,
je celý systém často, zejména v klinické literatuře, označován jako komplex mediálního spánkového laloku (medial temporal lobe;
MTL, Eichenbaum 2001). Úloha hipokampu v paměti je oceňována především od popisu případu pacienta Henry Gustava Molaisona
(dnes již zesnulý slavný „HM patient“), který utrpěl hlubokou anterográdní amnézii deklarativní,
především epizodické paměti po neurochirurgické resekci velké části mediálního spánkovém laloku včetně
hipokampu provedené terapeuticky pro farmakorezistentní temporální epilepsii (Scoville a Milner, 1957). Specifická teorie funkce hipokampu navrhuje,
že tato struktura se souvisejícími oblastmi jsou sídlem "kognitivní mapy" prostředí, zvláště užitečné při navigaci
ke skrytým cílům, tj. "navigace mapou" (Tolman 1948, O'Keefe a Nadel 1978). Tento vlivný koncept hipokampu dlouho poskytoval
hlavní rámec výzkumu této struktury, ale také bylo prokázáno, že hipokampus hraje klíčovou roli v úzkosti
(zejména jeho ventrální část; Bannerman et al 2004, Kheirbek et al 2013), v organizaci více proudů
informací (kognitivní koordinace; Wesierska et al 2005) a v kognitivní flexibilitě (Burghardt et al., 2012).
Hipokampus je také velmi důležitý pro rozpoznání pozice v prostoru i bez navigačních požadavků
(Klement et al. 2005) a pro rozpoznání polohy objektů v nepřístupném prostředí (Levčík et al. 2013).
Nedávno bylo navrženo, že hipokampus by mohl hrát roli spíše v prostorové volbě (či akci) než prostorové znalosti (Bannerman et al. 2012), což významně podtrhuje roli neokortexu.
Hippokampus je fylogeneticky stará mozková kůra, tzv. archikortex. Je propojen v širší funkční síť
sestávající z hippocampus proper (Cornu Ammonis, CA1-4), gyrus dentatus (angl. dentate gyrus; DG), and subikulárního komplexu (Amaral a Witter 1989).
Spolu s okolními oblastmi kůry, především entorhinální, perirhinální a postrhinální kůry,
je celý systém často, zejména v klinické literatuře, označován jako komplex mediálního spánkového laloku (medial temporal lobe;
MTL, Eichenbaum 2001). Úloha hipokampu v paměti je oceňována především od popisu případu pacienta Henry Gustava Molaisona
(dnes již zesnulý slavný „HM patient“), který utrpěl hlubokou anterográdní amnézii deklarativní,
především epizodické paměti po neurochirurgické resekci velké části mediálního spánkovém laloku včetně
hipokampu provedené terapeuticky pro farmakorezistentní temporální epilepsii (Scoville a Milner, 1957). Specifická teorie funkce hipokampu navrhuje,
že tato struktura se souvisejícími oblastmi jsou sídlem "kognitivní mapy" prostředí, zvláště užitečné při navigaci
ke skrytým cílům, tj. "navigace mapou" (Tolman 1948, O'Keefe a Nadel 1978). Tento vlivný koncept hipokampu dlouho poskytoval
hlavní rámec výzkumu této struktury, ale také bylo prokázáno, že hipokampus hraje klíčovou roli v úzkosti
(zejména jeho ventrální část; Bannerman et al 2004, Kheirbek et al 2013), v organizaci více proudů
informací (kognitivní koordinace; Wesierska et al 2005) a v kognitivní flexibilitě (Burghardt et al., 2012).
Hipokampus je také velmi důležitý pro rozpoznání pozice v prostoru i bez navigačních požadavků
(Klement et al. 2005) a pro rozpoznání polohy objektů v nepřístupném prostředí (Levčík et al. 2013).
Nedávno bylo navrženo, že hipokampus by mohl hrát roli spíše v prostorové volbě (či akci) než prostorové znalosti (Bannerman et al. 2012), což významně podtrhuje roli neokortexu.
nahoru
Neokortikální oblasti, prostorově specifické buňky a jejich možné role
 Několik neokortikálních oblastí propojených s hipokampem se účastní prostorové navigace, včetně např.
retrospleniální kůry. Retrospleniální kůra (angl. retrosplenial cortex; RSC) tvoří jakési funkční "rozhraní"
mezi hipokampem a neokortikálními asociačními oblasti a může hrát roli v integraci egocentrických a allocentrických proudů
informace (Byrne et al. 2007). Narušení RSC zhoršuje navigaci v úloze disociovaných rámců
(aktivní vyhýbání se místu na kolotočové aréně) (Wesierska et al. 2009), což ukazuje na její roli v kognitivní koordinaci.
Další oblastí, zapojenou do navigace a prostorové paměti je zadní parietální kůra,
která je zodpovědná především za egocentrické zpracování (Whitlock et al. 2012, ale viz Svoboda et al. 2009).
Význam entorhinální kůry je třeba rovněž zdůraznit, vzhledem k tomu, že poskytuje hlavní excitační
a informační vstup do gyrus dentatus a je vstupní branou do hipokampu .Důležité je, že neurony v rámci širší
limbické sítě u hlodavců vykazují různé typy prostorové aktivity. Pyramidové neurony v hipokampu se chovají jako buňky místa
(angl. place cells), tj. významně zvyšují vzruchovou aktivitu, pokud se subjekt nachází v definované a omezené části prostoru,
označovaného anglickým termínem place (či firing field) (O'Keefe a Dostrovsky 1971, přehled viz Muller, 1996).
Ostatní funkčně specializované buněčné typy jsou buňky směru hlavy (angl. head direction cells;
Taube et al., 1990) v subikulu, RSC a předním thalamu, mřížkové buňky (grid cells)
v mediální entorhinální kůře (Fyhn et al. 2004, Hafting et al. 2005) a buňky se smíšenými preferencemi
na směr hlavy a zároveň polohu (Cho a Sharp, 2001). Buňky směr hlavy reagují, když zvíře natočí
hlavu v určitém směru nezávisle na poloze v bludišti, zatímco mřížkové buňky vytvářejí vzruchy na mnoha místech,
která pravidelné pokrývají celý prostor hexagonální mřížkou. Jiné typy buněk reagují na prostorové hranice (border cells),
nebo na kombinace prostorových proměnných, jako je místo, směr hlavy, a rychlost pohybu.
Několik neokortikálních oblastí propojených s hipokampem se účastní prostorové navigace, včetně např.
retrospleniální kůry. Retrospleniální kůra (angl. retrosplenial cortex; RSC) tvoří jakési funkční "rozhraní"
mezi hipokampem a neokortikálními asociačními oblasti a může hrát roli v integraci egocentrických a allocentrických proudů
informace (Byrne et al. 2007). Narušení RSC zhoršuje navigaci v úloze disociovaných rámců
(aktivní vyhýbání se místu na kolotočové aréně) (Wesierska et al. 2009), což ukazuje na její roli v kognitivní koordinaci.
Další oblastí, zapojenou do navigace a prostorové paměti je zadní parietální kůra,
která je zodpovědná především za egocentrické zpracování (Whitlock et al. 2012, ale viz Svoboda et al. 2009).
Význam entorhinální kůry je třeba rovněž zdůraznit, vzhledem k tomu, že poskytuje hlavní excitační
a informační vstup do gyrus dentatus a je vstupní branou do hipokampu .Důležité je, že neurony v rámci širší
limbické sítě u hlodavců vykazují různé typy prostorové aktivity. Pyramidové neurony v hipokampu se chovají jako buňky místa
(angl. place cells), tj. významně zvyšují vzruchovou aktivitu, pokud se subjekt nachází v definované a omezené části prostoru,
označovaného anglickým termínem place (či firing field) (O'Keefe a Dostrovsky 1971, přehled viz Muller, 1996).
Ostatní funkčně specializované buněčné typy jsou buňky směru hlavy (angl. head direction cells;
Taube et al., 1990) v subikulu, RSC a předním thalamu, mřížkové buňky (grid cells)
v mediální entorhinální kůře (Fyhn et al. 2004, Hafting et al. 2005) a buňky se smíšenými preferencemi
na směr hlavy a zároveň polohu (Cho a Sharp, 2001). Buňky směr hlavy reagují, když zvíře natočí
hlavu v určitém směru nezávisle na poloze v bludišti, zatímco mřížkové buňky vytvářejí vzruchy na mnoha místech,
která pravidelné pokrývají celý prostor hexagonální mřížkou. Jiné typy buněk reagují na prostorové hranice (border cells),
nebo na kombinace prostorových proměnných, jako je místo, směr hlavy, a rychlost pohybu.
nahoru
Neurogeneze v dospělém hipokampu
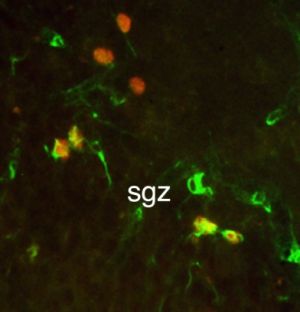 Důležité je také, že hipokampus, konkrétně DG, je jedním ze dvou míst neurogenezi v dospělém mozku (Altman a Das 1965).
Role nově zrozených neuronů jsou předmětem kontroverzí, některé teorie zdůrazňují
usnadnění separace podobných vzorců, tj. oddělení překrývajících se vzorů
v hipokampálních vstupech a tudíž i různých behaviorálních kontextech (Sahay et al. 2011, Nakashiba et al. 2012)
a zabránění rušení nových vzpomínek starými (tzv. prevence interference; Wiskott et al. 2006, Winocur et al. 2011).
Důležité je také, že hipokampus, konkrétně DG, je jedním ze dvou míst neurogenezi v dospělém mozku (Altman a Das 1965).
Role nově zrozených neuronů jsou předmětem kontroverzí, některé teorie zdůrazňují
usnadnění separace podobných vzorců, tj. oddělení překrývajících se vzorů
v hipokampálních vstupech a tudíž i různých behaviorálních kontextech (Sahay et al. 2011, Nakashiba et al. 2012)
a zabránění rušení nových vzpomínek starými (tzv. prevence interference; Wiskott et al. 2006, Winocur et al. 2011).
V poslední době jejich role byla přeformulována na zvýšení paměťového rozlišení (memory resolution),
kdy spolupráce mezi nově narozenými, hyperexcitabilním buňkami kódujících subtilní změny a starších neuronů,
které kódují výrazné rysy, zvyšuje množství detailů zakódovaných v mozkových reprezentacích (Aimone et al. 2011).
Nedávno byla také prokázána klíčová role neurogeneze v dospělosti pro udržení kognitivní flexibility, a a to za použití
rotující arény a blokády neurogeneze. Přes všechny dosavadní pokroky zůstávají role nově zrozených neuronů
v hipokampu z větší části nepopsané.
nahoru
Navigace v oddělených referenčních rámcích
Reálný svět, ve kterém žijeme, často obsahuje více referenčních rámců,
jakýchsi souřadnicových soustav, ve kterých můžeme určit svoji polohu (např. jedeme-li v autě,
existuje rámec okolní krajiny a rámec interiéru našeho vozu – a to je jen velmi triviální příklad).
Lidé stejně jako zvířata čelí denně také masivnímu proudu stimulů (informací), a to jak relevantních,
tak irrelevantních. Schopnost koordinovat mnohočetné vztažné rámce a spravovat vícečetné proudy informace je pro přežití zcela zásadní,
ale vyžaduje náročnou výpočetní kapacitu mozku. Většina studií zabývající se prostorovou orientací
jako modelem deklarativní paměti kupodivu dynamiku světa nabere příliš v potaz,
a navigace studuje často ve statickém uspořádání či po jednorázové změně prostředí,
ačkoliv existují i výjimky, kdy je cílem studia i dynamický aspekt (např. Ježek et al., 2012).
nahoru
Kognitivní deficity v animálním modelu
psychózy vyvolaném MK-801
Podání MK-801 potkanům či myším vytváří typické chování změny analogické k vybraným symptomům schizofrenie.
Navíc, MK-801 a další nonkompetitivní antagonisté NMDA receptorů mohou vyvolat akutní psychózu u
zdravých lidí a zhoršit příznaky u pacientů se schizofrenií (Newcomer a Krystal, 2011)
podporující fenomenologickou validitu těchto modelů založených na podávání antagonistů NMDA receptorů.
Model využitím MK-801 má kořeny v hypoglutamátergní hypotéze schizofrenie.
Ta předpokládá, že dysregulace glutamátergní neurotransmise je zásadním faktorem onemocnění.
V současnosti je kladen důraz i na neurovývojový aspekt a objevují se chronické a ontogenetické glutamátergní modely.
Akutní podání MK- 801 způsobuje dávkově závislou hyperlokomoce (vyšší dávky), sociální deficit (nízké dávky; Rung et al., 2005),
stereotypické chování a celkové ochuzení behaviorálního repertoáru (Nilsson et al., 2001).
Hyperlokomoce v modelech je poněkud „odvážně“ považována za analogii pozitivních symptomů u lidí;
tato analogie je založena na zvýšené dopaminové aktivitě v mesolimbických oblastech.
nahoru
V současnosti také rozvíjíme model obsedantně-kompulzivní poruchy (OCD) pomocí senzitizace D/D3 dopaminových receptorů quinpirolem. OCD je chronická a částečně dědičná porucha chování. Její celoživotní prevalence se odhaduje na 1-3 % (Stein 2002). OCD se vyznačuje opakujícími se dotěrnými myšlenkami (obsesemi) a opakujícími se ritualizovanými vzorci chování nazývanými kompulze, které často snižují úzkost vyvolanou obsesí (Stein 2002). OCD zasahuje pacienta na nejcitlivějších místech (myšlení, pohnutky, jednání, úzkost) a negativně ovlivňuje kvalitu života, a může dokonce zcela ovládat život postižených pacientů, kdy obsese a kompulze zabírají podstatnou část dne. Kromě toho značná část pacientů nereaguje na zavedené léčebné přístupy. První volbou léčby OCD jsou selektivní inhibitory zpětného vychytávání serotoninu (SSRI) a kognitivně-behaviorální terapie (KBT). V některých případech je také používání tricyklické antidepresivum klomipramin. Patofyziologie OCD je stále neznámá a onemocnění je, podobně jako schizofrenie, relativně heterogenní, především z hlediska obsesí a kompulzí. S rozvojem nemoci bývá spojována dysfunkce fronto-striatálních obvodů včetně orbitofronto-striato-thalamického systému. Nově byla tato teorie rozšířena o dorzolaterální prefronto-striatální systém spolu s parietální kůrou, což představuje podstatné rozšíření navrhovaného okruhu (Menzies et al. 2008, Kopřivová et al. 2009). Význačné a robustní změny jsou také nalézány v přední cingulární kůře (ACC) což je oblast mj. zodpovědná za detekci chyby a spekuluje se, že by v případě OCD mohla hrát roli v pacienty často zmiňovaném pocitu „něco není v pořádku“, který může představovat jakýsi spouštěč příznaků. A skutečně existuje teorie pojímající OC jako poruchu tzv. systému bezpečnostní motivace (security motivation system), kdy po vykonání behaviorální akce nedojde v nervovém systému k dostatečné zpětné vazbě, která by normálně tuto aktivitu ukončila.
nahoru